LA CELULA
Citoesqueleto:
 Es una estructura proteica que permite soportar la membrana plasmática, que mantiene la forma de la célula, facilita la movilidad celular (usando estructuras como los cilios y los flagelos), y desempeña un importante papel tanto en el tráfico intracelular, por ejemplo, los movimientos de vesículas y orgánulos y en la división celular.
Es una estructura proteica que permite soportar la membrana plasmática, que mantiene la forma de la célula, facilita la movilidad celular (usando estructuras como los cilios y los flagelos), y desempeña un importante papel tanto en el tráfico intracelular, por ejemplo, los movimientos de vesículas y orgánulos y en la división celular.
Las células eucariotas tienen tres tipos de filamentos cito esqueléticos: microfilamentos, filamentos intermedios y microtúbulos. Las septinas se consideran el cuarto componente del citoesqueleto.
Microfilamentos
citoesqueleto de actina:
- La actina es la proteína intracelular más abundante en eucariotas. Pueden llegar a representar un 10% en su peso total de proteína.
- Cada filamento está formado por dos cadenas entrelazadas en forma helicoidal, y cada cadena es un polímero de una proteína monomérica globular llamada actina.
- Puede alargarse en cualquiera de sus dos
extremos por el agregado de monómeros de actina “polimerización”
- La velocidad de crecimiento es mayor en uno de extremo (+) y el otro extremo (-)
- Pueden acortarse por la eliminación “despolimerización” de subunidades de actina a partir de cualquiera de sus extremos
- Si bien se extiende por todo el citoplasma
suelen acumularse en una capa justo por debajo de la membrana plasmática
formando una red que le confiere sustento, elasticidad y resistencia
mecánica
- Además, los monómeros de actina que se polimerizan poseen ATP y luego de ensamblarse en el filamento el ATP se hidroliza en ADP.

Microtúbulos y proteínas motoras:
- Son polímeros de la proteína tubulina que juega un papel muy importante en la división celular.
- Las proteínas globulares pueden también agruparse en diminutos túbulos huecos que actúan como entramado estructural de las células y al mismo tiempo transporta sustancia de una parte de la célula a otra.
- Están formados por dos tipos de moléculas
proteicas casi esféricas que se
disponen por parejas y se unen en los extremos creciente del microtúbulo y aumentan su longitud de las necesidades - Los microtúbulos constituyen también la estructura interna de los cilios y flagelos, apéndices de la membrana de los que se sirven algunos microorganismos para moverse.
- Existen proteínas que aprovechan la hidrólisis de ATP para generar energía mecánica y desplazar sustancias sobre microtúbulos. Estas son la dineína, transportador retrógrado, y la kinesina, transportador anterógrado.
- La dineína es una molécula de estructura similar a la kinesina: consta de dos cadenas pesadas idénticas que conforman dos cabezas globulares y de un número variable de cadenas intermedias y de cadenas ligeras.
- La mayoría de las kinesinas intervienen en el transporte anterógrado de vesículas, es decir, que implican un movimiento hacia la parte más distal de la célula o la neurita, desde el extremo (-) hacia el (+) de los microtúbulos, sobre los que se desplazan


Citoesqueleto y
movimiento celular
El
citoesqueleto proporciona una armazón estructural para la célula, actuando como
un andamio que determina la forma celular y la organización celular general del
citoplasma.
 Además
desempeña este papel estructural, el citoesqueleto es el responsable de los
movimientos de la célula, no solo incluye los movimientos en conjunto, sino
también el transporte interno de los orgánulos y otros y de otras estructuras
(cromosomas mitóticos) a través del citoplasma.
Además
desempeña este papel estructural, el citoesqueleto es el responsable de los
movimientos de la célula, no solo incluye los movimientos en conjunto, sino
también el transporte interno de los orgánulos y otros y de otras estructuras
(cromosomas mitóticos) a través del citoplasma.
El
citoesqueleto está constituido por tres tipos principales de filamentos de
proteína: Filamento de actina, filamentos intermedios y micro túbulos, que se
mantiene juntos y unidos a los orgánulos intracelulares y a la membrana
plasmática mediante varias proteínas accesorias.
Estructura y organización de los
filamentos de actina
- · La actina es la proteína más importante en el citoesqueleto y las proteínas que se unen a esta, son las que regulan las funciones del citoesqueleto, tales como movimiento y forma celular además de la organización en general del citoplasma
- · Los microfilamentos o filamentos de actina se encuentra principalmente alrededor de la membrana plasmática, donde actúan como soporte mecánico.
- · En células musculares, la actina constituye el 20% de la proteína celular.
- ·
Los mamíferos presentan al menos 6 genes
de actina diferentes, 4 en células musculares como lo son las presentes en
musculo cardiaco, estriado, liso vascular, y liso entérico. En estas la actina
presente es del tipo alfa, mientras que la beta se encuentra en celulares no
musculares.



BIOENERGÉTICA Y METABOLISMO
Mitocondrias, cloroplastos y peroxisomas
Los
orgánulos citoplasmáticos además de estar implicados en el transporte y
distribución de las proteínas:
Proporcionan
compartimientos especializados en los que tienen lugar diversas actividades
metabólicas. Una actividad fundamental de todas las células es generar energía metabólica
y son de los dos orgánulos que están dedicados específicamente al metabolismo
energético y a la producción de ATP.
Las
mitocondrias son responsables de generar la mayoría de la energía útil derivada
de la degradación de los lípidos y de los carbohidratos y los cloroplastos
utilizan la energía obtenida de la luz solar para generar tanto ATP a partir de
Co2 y H2O
El peroxisoma contiene enzimas que intervienen en diversas
rutas metabólicas incluyendo en la degradación de los ácidos grasos y el
metabolismo derivado de la fotosíntesis.
- las proteínas
destinadas a las mitocondrias, cloroplastos y peroxisomas, en lugar de
sintetizarse en los ribosomas unidos a membranas y ser trasladados al retículo
endoplásmico.


Organización
y función de las mitocondrias.
 Están rodeadas por un sistema de doble membrana,
constituido por una membrana mitocondrial interna y otra externa separadas por
un espacio intermembrana.
Están rodeadas por un sistema de doble membrana,
constituido por una membrana mitocondrial interna y otra externa separadas por
un espacio intermembrana.
·
Desempeñan un papel
crucial en la generación de energía metabólica de la célula
eucariota
·
La mayoría de las
proteínas mitocondriales son traducidas en los ribosomas citoplasmáticos libres
y son importadas al orgánulo debido a señales directoras especificas
·
Contienen su propio ADN
que codifica al ARNt, ARNr, y algunas proteínas mitocondriales.
·
La
membrana interna forma numerosos pliegues “crestas” que se extiende hacia el
interior “o matriz” del orgánulo.
·
Cada
uno desempeña un papel funcional distinto, siendo la matriz la membrana interna
los principales comportamientos funcionales de la mitocondria.
·
La
fuente principal de energía metabólica en las células animales es la
degradación oxidativa de la glucosa (glucolisis) tiene lugar en el citoplasma,
donde la glucosa es convertida a piruvato.
·
El
piruvato es posteriormente trasportado
al interior de la mitocondria, donde se oxida completa a CO2
produce la mayor parte de la energía es utilizable ATP
 ·
La oxidación de los ácidos grasos también produce acetil
CoA que da forma similar es metabolizado
por el ciclo del ácido cítrico en las mitocondrias
·
La oxidación de los ácidos grasos también produce acetil
CoA que da forma similar es metabolizado
por el ciclo del ácido cítrico en las mitocondrias
·
La
oxidación del acetil CoA a CO2 esta acoplado a la reducción de Nad y Fadh, fad respectivamente. Por lo tanto la
mayor parte de la energía derivada del metabolismo oxidativo.
·
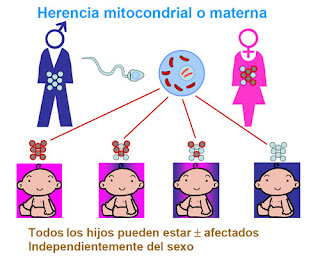
“Puesto
que casi todas las mitocondrias son heredadas de la madre, es posible determinar
el linaje materno humano hasta nuestra ancestro femenino común más reciente:
Eva mitocondrial”
Internalización
de proteínas y formación de las
mitocondrias
La mayoría de los genomas
mitocondriales no codifican las proteínas requeridas para la recopilación,
transcripción o traducción del ADN. Las proteínas requeridas para la
replicación y expresión del ADN mitocondrial se encuentra en el núcleo, contiene los genes
que codifican la mayoría de las proteínas mitocondriales requeridas para la fosforilación

Sistema
genético de la mitocondria
- Las mitocondrias contienen su propio sistema genético el cual esta separado y es distinto del genoma nuclear de la célula.
- Las mitocondrias son orgánulos subcelulares dedicados a la producción de energía en forma de ATP que contienen su propio sistema genético.
- El ADN mitocondrial codifica un número pequeño de polipéptidos que son indispensables para la vida celular por formar parte de la cadena respiratoria.
- Para la biogénesis de la mitocondria se requiere la expresión coordinada de los dos sistemas genéticos celulares, el nuclear y el mitocondrial. Los genes están dispuestos en el ADN mitocondrial de una forma extremadamente compacta con los ARNt intercalados entre los genes de los ARNr y los codificantes de proteínas.
- Esta organización génica se ve también reflejada en su modo de expresión y en las características singulares de los ARN. Las dos cadenas del ADN mitocondrial se transcriben completamente en forma de tres moléculas policistrónicas que se procesan posteriormente por enzimas específicas que cortan en los extremos 5 y 3 de los ARNt para originar los ARNr, ARNm y ARNt maduros.
- La genética del ADN mitocondrial difiere de la del ADN nuclear en una serie de propiedades. En particular, el genoma mitocondrial se hereda exclusivamente de la madre, que lo transmite a todos sus hijos. Este ADN tiene una tendencia a mutar más frecuentemente que el ADN nuclear por lo que se está utilizando para estudiar la filogenia y estructura de poblaciones.
- Clínicamente, hay un comienzo agudo de pérdida
visual, bilateral y simétrica. Esto finalmente evoluciona a una atrofia óptica muy
severa y una disminución permanente de agudeza
visual.







Síntesis de ATP
- la energía almacenada en el gradiente de protones a través de la membrana del tilacoides, es de naturaleza química casi en su totalidad. Debido que la membrana del tilacoide aunque es impermeable a los protones, difiere de la membrana mitocondrial interna en que es permeable a otros iones concretamente MG2 y al Cl
- sin embargo puesto que la luz del tilacoide es un comportamiento cerrado esta diferencia en la concentración de protones puede ser bastante grande.
- El gradiente de protones se produce como resultado de la entrada de NADH (producido en las reacciones de oxido-reducción ) a la cadena transportadora de electrones . Los protones se acumulan en el espacio intermembrana hasta un gradiente de concentración tal que pueden ser utilizados para producir ATP.

2. ) Los Protones (indicados
por +) entran nuevamente en la matriz mitocondrial a través de los canales
que forma el complejo enzimático de la ATP sintetasa. Esta entrada
se acopla a la síntesis de ATP a partir de ADP y Fosfato (Pi)

Peroxisomas
Son
orgánulos pequeños, rodeados por unas membranas que contienen enzimas
implicadas en diversas reacciones metabólicas, incluyendo varios aspectos del
metabolismo energético
- · la mayoría de las células humanas contienen unos 500 peroxisomas.
- · Los peroxisomas no poseen su propio genoma humano que codifican proteínas peroxisomales, la cual en su mayoría de las cuales actúan como enzimas metabólicas.
- · La mayoría son sintetizadas sobre ribosomas libres y después importadas en los peroxisomas como cadenas polipeptídicas completas.
·
Entre otras funciones de los peroxisomas, estos también se encargan de desencadenar reacciones
oxidativas. Esto no proporciona energía en forma de ATP, pero
permite producir calor, teniendo gran importancia fisiológica.
·
La formación de los peroxisomas se
lleva a cabo en el retículo, y tienen la capacidad de dividirse gracias a su
crecimiento y
Estrangulamiento,
lo cual ocurre generalmente durante la división celular. Este proceso es
llevado a cabo por el citoesqueleto y proteínas.

Núcleo
La existencia del núcleo es la característica principal:
 Que
diferencia las células procariotas, por contener el genoma celular, el núcleo
sirve de almacén de la información genética y como centro de control celular,
la recopilación del ADN, la transpiración y el procesamiento del ARN ocurre en
el interior del núcleo, y solo la última etapa de la expresión genética tiene
lugar en el citoplasma.
Que
diferencia las células procariotas, por contener el genoma celular, el núcleo
sirve de almacén de la información genética y como centro de control celular,
la recopilación del ADN, la transpiración y el procesamiento del ARN ocurre en
el interior del núcleo, y solo la última etapa de la expresión genética tiene
lugar en el citoplasma.
Debido
a que la envuelta nuclear separa el genoma del citoplasma, la expresión génica está
regulada por mecanismos exclusivos de los organismos eucariotas
·
Mientras que los ARNm
procariotas son traducidos a la vez que ocurre la transpiración
·
Ejemplo: la expresión
de algunos genes eucariotas se controla a través de la regulación
transcripcional inexistente en procariotas.
Envuelta nuclear y tráfico entre el núcleo
y el citoplasma
·
La envuelta nuclear
separa el contenido del núcleo del citoplasma y proporciona una armazón
estructural al núcleo.
·
Las membranas nucleares
actúan como una barrera selectiva que impide el libre paso de las moléculas
entre el interior del nuclear y citoplasma, manteniéndolos como dos
comportamientos metabólicamente independientes.
·
Los únicos canales
están representados por los complejos de poros nucleares que permiten un
intercambio controlado de moléculas entre el núcleo y el citoplasma.
·
El tráfico selectivo de
proteínas y ARN a través de los complejos de poro nucleares no solo mantiene la
composición interna del núcleo sino que tiene un papel clave en la regulación
de la expresión genética.

 |
 |
| Añadir leyenda |
Estructura de la envuelta celular.
Posee una estructura compleja,
constituida por dos membranas nucleares, la lámina nuclear en su cara interna y
por los complejos de los poros nucleares.
El núcleo está delimitado por
un sistema de dos membranas concéntricas
- · Membrana nuclear interna y externa.
- · La externa se continúa con la membrana del retículo endoplasmático, por lo que hay una comunicación directa entre el espacio intermembrana y el lumen del retículo endoplasmático, posee ribosomas adheridos a su superficie citoplasmática si bien su composición proteica es ligeramente diferencial ya que es rica en proteínas de membrana que se une al citoesqueleto y carece de las proteínas que mantiene la org, cilíndrica del RE.
- · La membrana nuclear interna tiene proteínas únicas que son específicas para el núcleo como aquellas que unen la matriz nuclear de láminas
Su función principal es actuar
como una barrera que separa el contenido del interior del nuclear del
citoplasma.
La membrana celular es una bicapa fosfolipídica, la membrana nuclear externa e interna se unen en los complejos de poro nuclear
siendo los únicos canales que permiten el paso de pequeñas moléculas polares
macromoléculas a través de la envuelta nuclear.
- En la membrana nuclear interna se localiza la lamina nuclear una red fibrosa que proporciona soporte estructural al núcleo






Complejo del poro nuclear
Son los únicos canales a
través de los cuales pueden viajar pequeñas moléculas polares, iones y
macromoléculas (proteínas y ARN) entre el nucleo y el citoplasma.
·
El complejo del poro
nuclear es una estructura muy grande con un diámetro de aproximadamente 120 nm
y un peso moléculas estimado de a
proximidad 125 millones de daltons 30 veces el tamaño de un ribosoma
· Las moléculas de ARN que son sintetizadas en el núcleo
deben ser exportadas de manera eficiente al citoplasma donde intervienen en la
síntesis de proteínas

Transporte selectivo de proteínas desde y hacia el núcleo
·
El mecanismo del tráfico
selectivo a través de la envuelta nuclear se encuentra mejor caracterizado en
el caso de las proteínas que son importadas desde el citoplasma del núcleo.
·
Estas proteínas son
responsables de todas las características de la estructura y de la función del
genoma incluyen las histonas, ADN polimerasa, ARN polimerasas, factores de
transpiración, factores de splicing y otras más.

DISTRIBUCIÓN Y TRANSPORTE DE PROTEÍNAS
Retículo endoplásmico, aparato de Golgi y lisosomas
Además de por la presencia de un núcleo, las células eucariotas se distingue de las células
procariotas por la presencia en el citoplasma de orgánulos rodeados de
membrana.
·
El primer paso en la
distribución de las proteínas tiene lugar mientras aún está en marcha la
traducción. Muchas proteínas destinadas al retículo endoplasmático, al aparato
de Golgi, a los lisosomas, a la membrana plasmática y a ser secretadas se
sintetizan en los ribosomas unidos a la membrana del retículo endoplásmico.
·
Desde el retículo endoplásmico, las
proteínas se transportan en vesículas al aparato de Golgi, donde son nuevamente
procesadas y distribuidas para el transporte a los lisosomas, a la membrana
plasmática o al ser secretadas desde la célula.
Retículo
endoplásmico:
·
Es una red de túbulos y
sacros “cisternas” rodeadas de membrana que se extiende desde la membrana
nuclear por todo el citoplasma.
·
Sus membranas se continúan con las del núcleo y se pueden extender hasta las proximidades
de la membrana plasmática, llegando a representar más de la mitad de las
membranas de una célula.
·
Todo el R.E está rodeado por una membrana continua y
es el orgánulo más grande de la mayoría de las celulares eucariotas.
Como se trato previamente hay dos tipos
distintos de r.e que realizan funciones diferentes en la celula:
·
El rugoso, que posee membranas
formando túbulos más o menos rectos, a veces cisternas aplanadas, y con numerosos
ribosomas asociados, y
- El liso, sin ribosomas asociados y con membranas organizadas formando túbulos muy curvados e irregulares


Comentarios
Publicar un comentario